Floral structures
traditionally hold the defining aspects of species. There are numerous characters associated with them. Basically there are two contrasting forms of floral architecture, the individual flower buds or flowers, and then their arrangement on the branchlets.
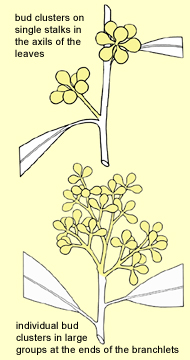
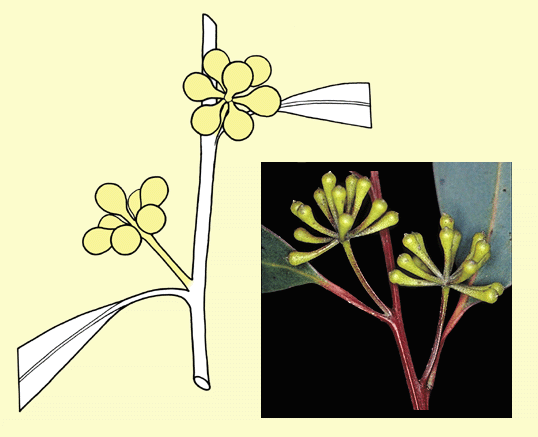
In most species of eucalypts, the buds occur
in
clusters on single stalks in the axils of the leaves. The flowers
are mostly small and whitish and are not conspicuous in the crown.

A very few species have the inflorescences in complex clusters in the leaf axils, e.g. E. michaeliana or on expanded axillary shoots as in some ghost gums, e.g. C. bella, C. tessellaris, and the spotted gums e.g. C. henryi, C. maculata, or in more contracted though still branched axillary shoots as in most ghost gums e.g. C. flavescens, C. polysciada. Four species from eastern Australia, E. fastigata, E. pachycalyx, E. regnans and E. squamosa, form their buds consistently in twin clusters in
the leaf axils. In contrast, several large groups, the bloodwoods, some
of the boxes and some of the ironbarks, form the individual bud clusters in large groups at the esnds of the branchlet, with
few or no leaves. In season these result in conspicuous sprays of flowers
on the outside of the crown. A prominent example is the yellow bloodwood
(C. eximia) of the sandstone regions of central eastern New South
Wales, where the creamy white flower clusters stand out in the forest.
In the south-west of Western Australia the widespread marri (C. calophylla) exhibits the same prolific flowering affect, although
the southern Red-flowering gum (C. ficifolia) and the commonly cultivated northern Swamp bloodwood,
C. ptychocarpa, are the most spectacular of the flowering eucalypts.
One species, E. cladocalyx, has ramiflorous inflorescences, with the buds formed
on the leafless part of the branchlets well inside the crown. Some ghost
gums from northern Australia which are deciduous in the dry season,
e.g. C. confertiflora, also appear to flower on leafless
branches but these are cases where the floral buds have formed in the
axils where last-season’s leaves used to be and the inflorescences
are axillary, not truly ramiflorous. Very useful diagnostic information
can be derived from these inflorescence patterns, although the structures
can be modified by various external factors including predation.
A common modification of the basic axillary inflorescence of the eucalypts
can be seen in many 'box', 'ironbark' and 'bloodwood' species. In these,
bud clusters are formed in the usual way in the axils of developing
leaves towards the ends of the annual growth of a branchlet. The arrangement
of these leaves and floral primordia is initially decussate, and subsequent
uneven elongation of the axis gives the appearance of alternation. Each
branchlet terminates with a vegetative bud. In many 'box', 'ironbark'
and 'bloodwood' species, this terminal vegetative bud aborts and the
now apparently alternate leaf primordia cease their development. The
floral primordia however, continue to develop, resulting in a 'leafless'
compound inflorescence, terminating the branchlet. Good examples of
this are E. paniculata, the common grey ironbark of south-eastern Australia,
and C. calophylla,
or Marri, common in south-western Australia.

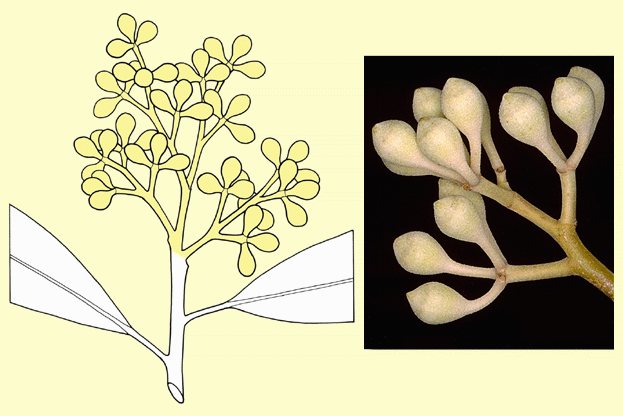
The individual bud clusters in most eucalypts can be seen on close
inspection to be in symmetrical patterns. A few species have a single
bud in the inflorescence, e.g. E. globulus and E. macrocarpa, but the basic numbers in Angophora, Corymbia
and Eucalyptus are 3 or 7. In a 3-budded inflorescence
there is a central erect bud and two subtending side buds, all in a
plane at right angles to the stem, forming a 'cross'. A 7-budded inflorescence
has a central erect bud, two subtending side buds plus two buds each
subtending the side buds. Bud numbers higher than 7 form by the addition
of further pairs of subtending buds, and the number of buds in an intact
inflorescence is always odd (never an even number), although very high
bud numbers may occur in an obscured pattern. Also, in inflorescences
with high numbers, one of a pair of subtending buds may be suppressed,
probably by compression in the very young inflorescence which is tightly
held within bracts
which are soon deciduous. When assessing bud numbers, it is important
to take into account the fact that during inflorescence development,
which often takes more than a year, individual buds may be lost. This
is particularly the case by the fruiting stage when the structures under
examination have been exposed for a long time and subject to various
traumas including predation and simple death of individual buds.
Angophora species and some of the northern bloodwoods (Corymbia setosa and related species) have simple hairs and bristle glands
(erect multicellular hairs or setae) somewhere on the inflorescence,
peduncle, pedicel, and often on the bud. The buds of Eucalyptus
species are glabrous for their whole life cycle.
 Angophora
Angophora flowers

Inner opercula
Operculum scars
Inner bud anatomy
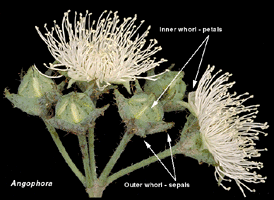
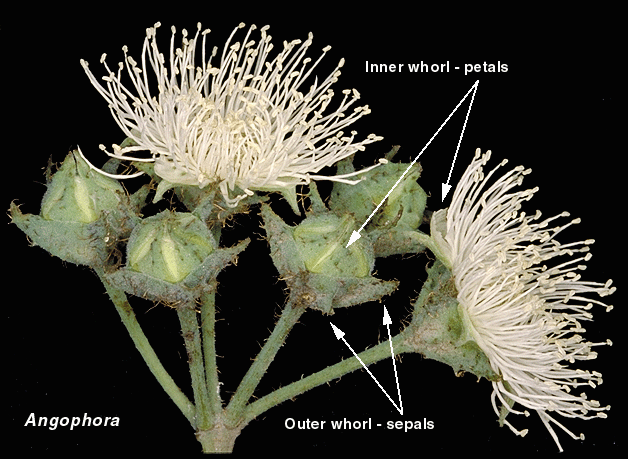
Angophora species are readily distinguished from other eucalypts
in the flowers, by the presence of petals that have a green keel and
white margin, and by persistent hard, woody, green sepals.
All Corymbia species and most Eucalyptus species do
not have separate sepals. The exceptions are the species in Eucalyptus
subgenus Eudesmia plus a handful of other species. Subgenus
Eudesmia is widespread and consists of 21 species. In south-western
Western Australia the most famous is the glaucous, juvenile-leaved Tallerack
(E. pleurocarpa). In this and related species, the calyx is formed
of distinct separate sepals which are usually evident as four small teeth at the top of the hypanthium and usually persist to the fruiting stage. A northern example is the Darwin Stringybark,
E. tetrodonta, which in bud has prominent sepals that persist
in fruit. Another group of eudesmids have their sepals more or less fused to the corolla right at the apex of the bud and
usually are difficult to see. Examples of this are E. baileyana from Queensland and northern New South Wales, E. ebbanoensis
from south-western Western Australia, and the orange-flowered tropical
trees E. miniata and E. phoenicea.
Other Eucalyptus species having separate sepals are E.
microcorys, which has, in early bud development, very small calyx
lobes formed at the top of the hypanthium but which fall early and are
seldom seen, and the south-western species E. steedmanii and
E. mimica where conspicuous sepals are present in bud but are
lost on flowering; the Queensland endemic species E. curtisii,
E. cloeziana and E. tenuipes, with four small teeth present
on the mid line of the bud which persist in E. curtisii but fall
early in the other two. In all other species in Eucalyptus and
in Corymbia the sepals are united to form the outer operculum
or bud-cap.

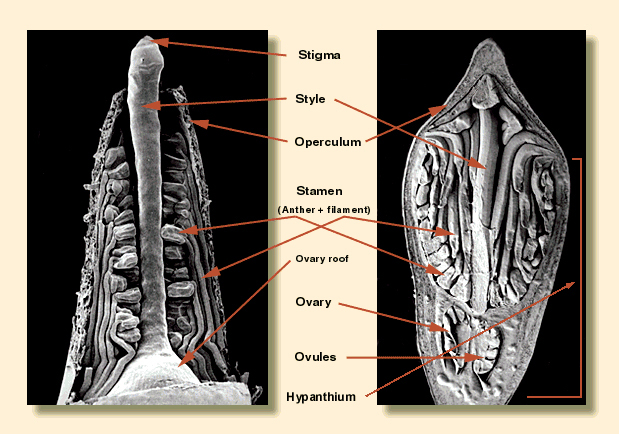
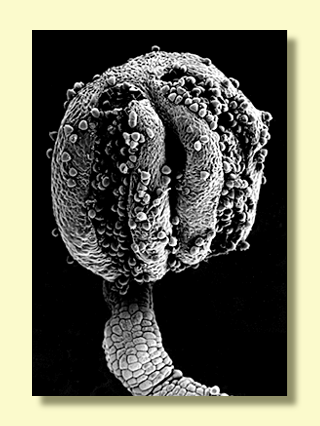
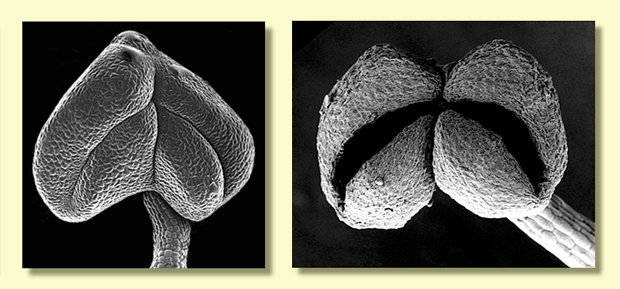
The individual flower buds have two opercula (bud caps covering the
stamens and style) derived from the united sepals (outer operculum)
and united petals (inner operculum). In some species of red bloodwood
the fusion of the petals to form the inner operculum may not be complete,
but careful dissection is needed to see this. A longitudinal section
through an almost mature bud can reveal whether or not the inner operculum
is divided at all. Similarly, removing the outer operculum but leaving
the inner operculum intact can also show whether the inner operculum
is partially divided or not. Some examples in the bloodwoods are C.
ficifolia, C. zygophylla and C. deserticola. Eucalyptus
guilfoylei from the wet forests of southern Western Australia may
also possess this feature of the inner operculum.
The flower buds of Angophora (illustrated above) are all very
similar within the group of twelve species and subspecies and, apart
from size, contain very few discernible characters that distinguish
the species. The individual flower buds of the traditional eucalypts,
however, contain a great deal of vital information, from the external
superficial nature of the wall of the bud to the characters of much
higher reliability contained within. One character of absolute reliability
(no exceptions have ever been found) is the number of opercula, although
this requires experience to assess.

Except for Angophora, the eucalypt flower lacks showy petals. The
petals are in fact united very early in bud development to form a cap
or a cone-shaped structure that covers the stamens and ovary during
their development. This is the inner operculum, which sheds just before flowering when the stamens
expand and are almost ready to shed their pollen. (There is a delay
in pollen ripening and dispersal to lessen the chance of self-fertilisation
and consequent inbreeding). The outer whorl of the floral parts is the
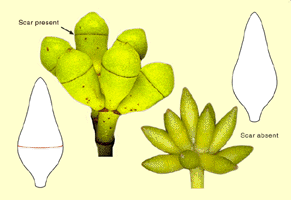
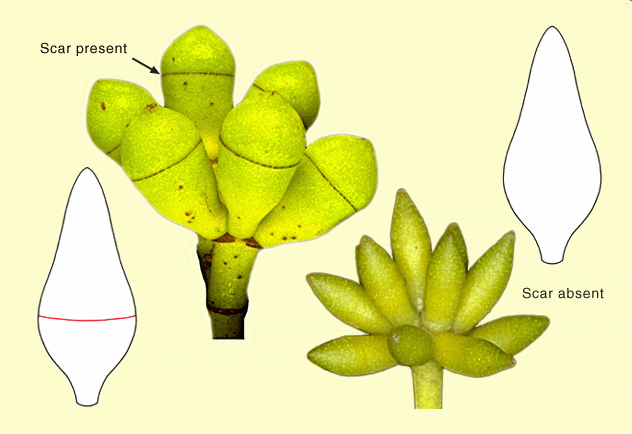
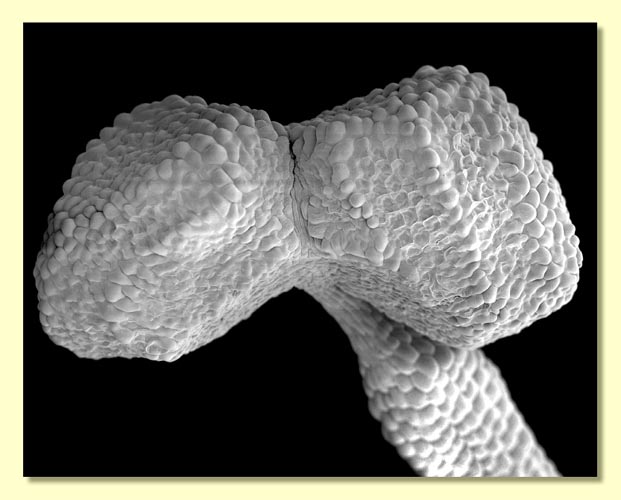
sepals which, likewise, unite to form an operculum in most eucalypt
species. In the majority of species, this, the outer operculum sheds
early in bud development. In doing so, the tissue around the approximate
middle of the bud, i.e. where the outer operculum attaches to the base
of the bud, dies resulting in detachment. This leaves a scar around the middle of the bud which can sometimes be seen with
the naked eye but is best seen with a lens. About 130 species, comprising
the Eucalyptus subgenus Eucalyptus, have lost the
outer operculum altogether in the evolution of the group. Therefore,
throughout the development of the bud in these species there is no scar, and the side of the bud is smooth. Some species have two opercula
that are fused giving the superficial impression that only a single
operculum is present, e.g. E. ochrophloia. The boxes and ironbarks
show parallel development in operculum characters. There are two groups,
one in which the outer operculum sheds early leaving a scar, e.g. the
box species, E. behriana, and the ironbark species, E.
paniculata, and another in which the outer operculum is held to
bud maturity, e.g. the box species, E. microcarpa and the ironbark,
E. sideroxylon. The double opercula and their retention to
bud maturity is a diagnostic feature of all the red bloodwoods (Corymbia
informal section Rufaria). The ghost gums (Corymbia
informal section Blakearia e.g. C. bella) and spotted
gums (Corymbia informal section Politaria e.g.
C. citriodora) shed the outer operculum during bud development
leaving an operculum scar.

Stamens
have various forms of orientation in the unopened bud. Some species
have their stamens wholly erect. Others have them uniformly inflexed,
while others have irregular orientation. Again, the extremes of positioning,
i.e. complete inflexion or complete erection, are easy to assess. However
there will be 'in-between' species in which the character is difficult
to categorise. 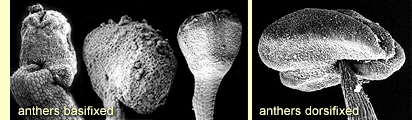 The
attachment of the anther on the summit of the staminal filament is useful
diagnostically. Some anthers are basifixed, with the tip of the filament
attached rigidly at the base of the anther. This character is seen in
the boxes and ironbarks and at its most extreme in E. leptophylla,
E. foecunda and related species. In the majority of eucalypts
the anthers are dorsifixed, by attachment loosely to the back of the
anther, such that it can swivel, i.e. versatile. Some eucalypts have
flowers with staminodes, where the outer stamens lack anthers or have
reduced, non-functional anthers, e.g. E. calycogona. The openings of the anther for pollen shed (dehiscence)
is also an important diagnostic character. Most eucalypts have their
anthers either opening by well separated longitudinal slits for the more or less cuboid anther, or, as in Eucalyptus
subgenus Eucalyptus (e.g. E. regnans) with their more
or less kidney-shaped anthers, have the openings oblique and touching
near the apex, finally forming confluent slits. The cuboid, freely dorsifixed anther occurs in many western
species but the kidney-shaped anther with confluent slits is rare in
western monocalypts but is seen in Jarrah (E. marginata) and
a few related species. The butterfly-shaped anther in E. guilfoylei is unique in the genus. In a considerable number
of species, particularly mallees, e.g. E. oleosa, the anthers
are subversatile and open by small roundish pores, either at the sides or the top of the anther.
The
attachment of the anther on the summit of the staminal filament is useful
diagnostically. Some anthers are basifixed, with the tip of the filament
attached rigidly at the base of the anther. This character is seen in
the boxes and ironbarks and at its most extreme in E. leptophylla,
E. foecunda and related species. In the majority of eucalypts
the anthers are dorsifixed, by attachment loosely to the back of the
anther, such that it can swivel, i.e. versatile. Some eucalypts have
flowers with staminodes, where the outer stamens lack anthers or have
reduced, non-functional anthers, e.g. E. calycogona. The openings of the anther for pollen shed (dehiscence)
is also an important diagnostic character. Most eucalypts have their
anthers either opening by well separated longitudinal slits for the more or less cuboid anther, or, as in Eucalyptus
subgenus Eucalyptus (e.g. E. regnans) with their more
or less kidney-shaped anthers, have the openings oblique and touching
near the apex, finally forming confluent slits. The cuboid, freely dorsifixed anther occurs in many western
species but the kidney-shaped anther with confluent slits is rare in
western monocalypts but is seen in Jarrah (E. marginata) and
a few related species. The butterfly-shaped anther in E. guilfoylei is unique in the genus. In a considerable number
of species, particularly mallees, e.g. E. oleosa, the anthers
are subversatile and open by small roundish pores, either at the sides or the top of the anther.
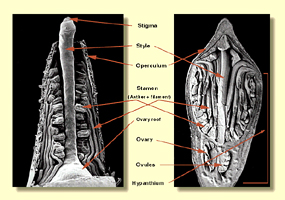
Within the base of the bud is the ovary and this contains characters of
high diagnostic reliability. The most useful is the number of vertical
rows of ovules.
These can only be seen by dissection and is best done under a microscope
but can be done in the field and seen with a 10× lens. Most eucalypts
have ovule rows with 4 or 6 vertical
rows. Another group has ovule rows consistently in 2s (Eucalyptus subgenus Eucalyptus),
while others have rows of 3
or 5, or irregular patterns (bloodwoods and ghost gums).

The top of the ovary is surmounted by the style which terminates in the
stigma. The style is usually erect in all but a few species but can
be spiral in some e.g. E. albida, making it a useful diagnostic
character. In the great majority of species the style arises from the
narrowed summit of the ovary. In some bloodwoods, in Eucalyptus series Melliodorae (e.g. E.
leucoxylon) and some species of Eucalyptus series Loxophlebae (e.g. E.
loxophleba) the style narrows at the base and is inserted into the roof
of the ovary. The style is subsequently articulate, not rigid.
The pollen is transported to the stigma from another flower by insects,
small birds or small mammals. On germination of the pollen grains, the
contents including the vital nuclei migrate by means of a pollen tube
down the stigma shaft to the ovary itself where several ovules at the
base of the placentae are fertilised. The fertilised ovules mature into
the seeds. The ovular structures on the upper part of the placentae
are infertile or unfertilised and 'mature' into sterile particles smaller
than the seeds known as the chaff.


 The
'internal' features of the eucalypt plant, such as the number of opercula
in the bud, arrangement of stamens, number of ovule rows and seed shape,
are usually more reliable for identification than the 'external' features.
They are relatively protected from the elements and from various forms
of predation. They are the parts that require handling and close inspection
or even dissection, as opposed to macro observation.
The
'internal' features of the eucalypt plant, such as the number of opercula
in the bud, arrangement of stamens, number of ovule rows and seed shape,
are usually more reliable for identification than the 'external' features.
They are relatively protected from the elements and from various forms
of predation. They are the parts that require handling and close inspection
or even dissection, as opposed to macro observation. 



 The
irregular markings on the living bark of some smooth bark species are
known as scribbles and are caused by burrowing insect larvae. Insects
are attracted to some species and not others, whether to eat the leaves,
suck nectar or to lay their eggs. Some insects are particularly partial
to species in Eucalyptus subgenus Eucalyptus - stringybarks,
ashes, peppermints, and related species, and lay their eggs in the bark.
The larvae then eat their way through the surface of the bark leaving
a characteristic zig-zag trail or scribble.
The
irregular markings on the living bark of some smooth bark species are
known as scribbles and are caused by burrowing insect larvae. Insects
are attracted to some species and not others, whether to eat the leaves,
suck nectar or to lay their eggs. Some insects are particularly partial
to species in Eucalyptus subgenus Eucalyptus - stringybarks,
ashes, peppermints, and related species, and lay their eggs in the bark.
The larvae then eat their way through the surface of the bark leaving
a characteristic zig-zag trail or scribble. 
 The
ironbarks only occur in northern and eastern Australia but some species
from south-western Western Australia have very hard rough bark that
is thinner than that of the eastern ironbarks to which they are only
very distantly related, e.g.
The
ironbarks only occur in northern and eastern Australia but some species
from south-western Western Australia have very hard rough bark that
is thinner than that of the eastern ironbarks to which they are only
very distantly related, e.g. 


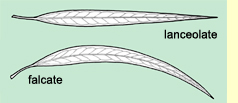 Adult
leaf shape is not much use in identification as most species have lanceolate
or falcate (curved) leaves. Leaf shape is a character of low reliability
for identification. Leaf size is less useful as many species have leaves
about the same size. It is most useful if the species typically has
adult leaves much larger (e.g. E. globulus) or much smaller
(e.g. E. parvula) than most other species.
Adult
leaf shape is not much use in identification as most species have lanceolate
or falcate (curved) leaves. Leaf shape is a character of low reliability
for identification. Leaf size is less useful as many species have leaves
about the same size. It is most useful if the species typically has
adult leaves much larger (e.g. E. globulus) or much smaller
(e.g. E. parvula) than most other species.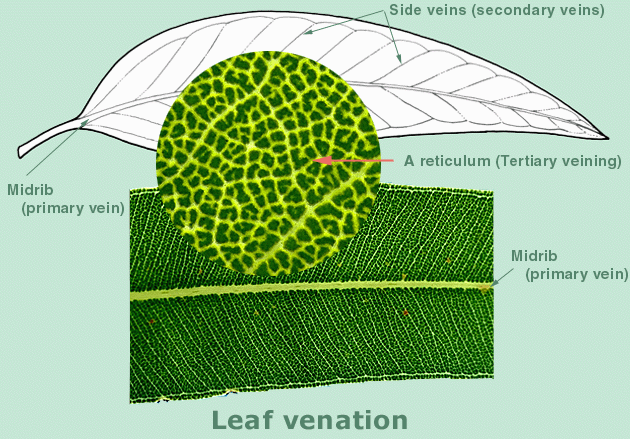




















 Once
a specimen has been taken, a very handy and accessible feature is the
pith of
the branchlets. In the southern half of the country about half of the
dry country mallees have a line of clear-coloured or
Once
a specimen has been taken, a very handy and accessible feature is the
pith of
the branchlets. In the southern half of the country about half of the
dry country mallees have a line of clear-coloured or {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}